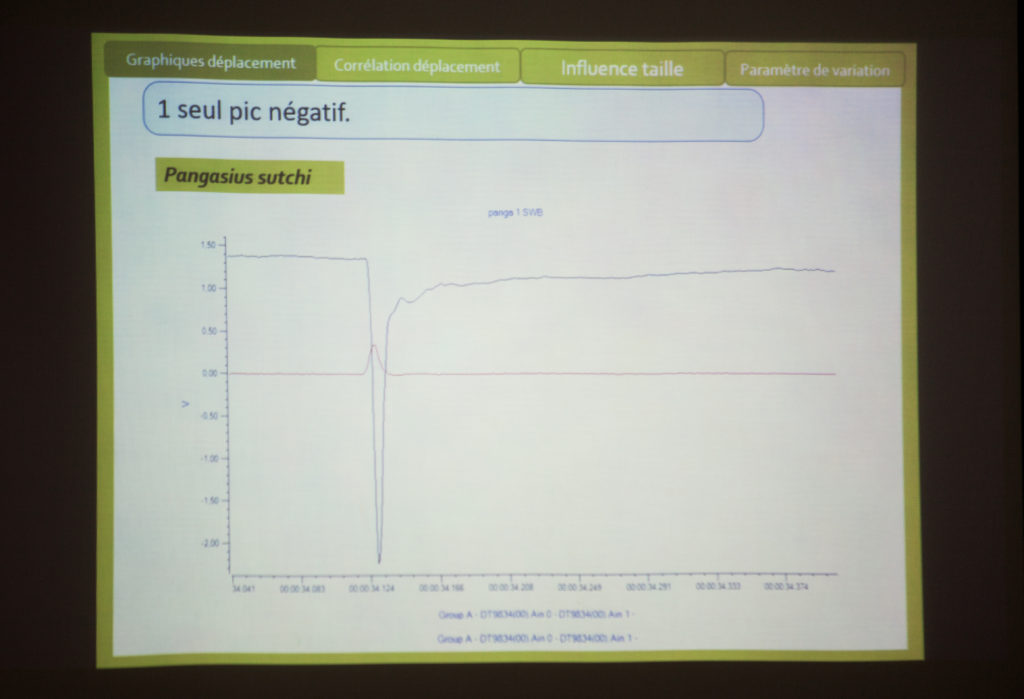
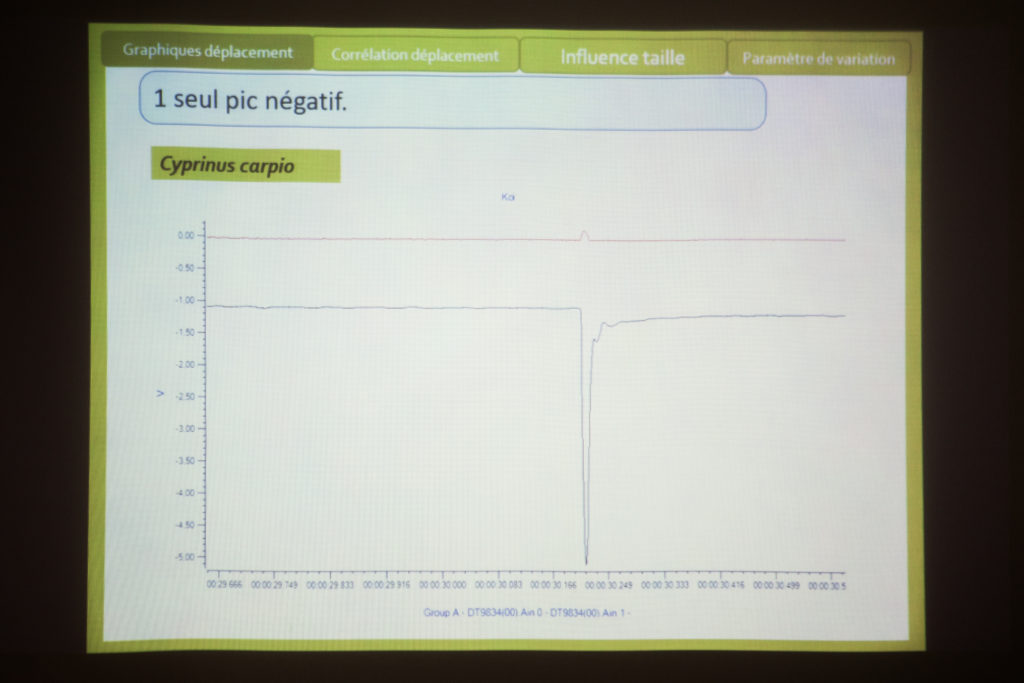
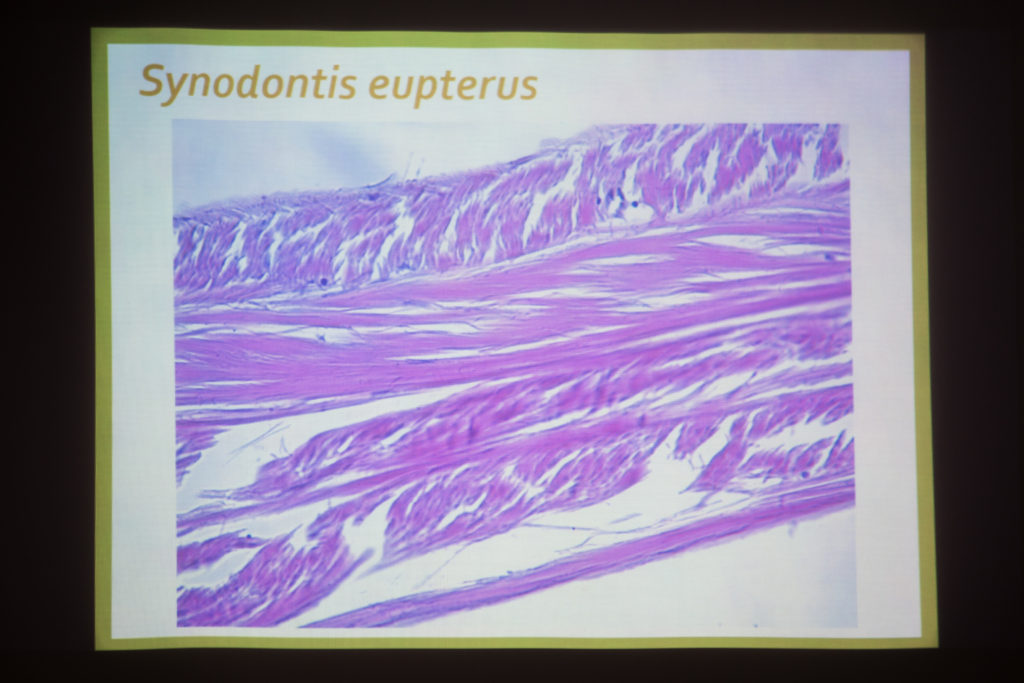
Écoute de quelques chants de poissons (maigre, mérou, carapidés) avec Eric Parmentier et discussion sur l’anatomie des poissons. Un exemple particulier : le carapus . Eric : « voilà le crâne, la colonne vertébrale, les côtes. La côte qui est un peu transformée, en forme de cuillère. Ça se sont les muscles qui permettre d’étendre la vessie. Peut-être pour faire varier, comme on le disait, en fonction de la profondeur. Les muscles sont tout à fait particuliers : ils sont longs et viennent se loger en dessous de la cuillère, le muscle a une épaisseur plus fine. ». Eric note que cela reste une vue d’artiste. La partie avant est tirée par le muscle puis revient et cogne, lorsque le muscle est relâché. « cogner ou faire bouger les côtes… on sais pas trop ! Ou alors, c’est la cuillère avec la paroi juste en dessous… voilà.»
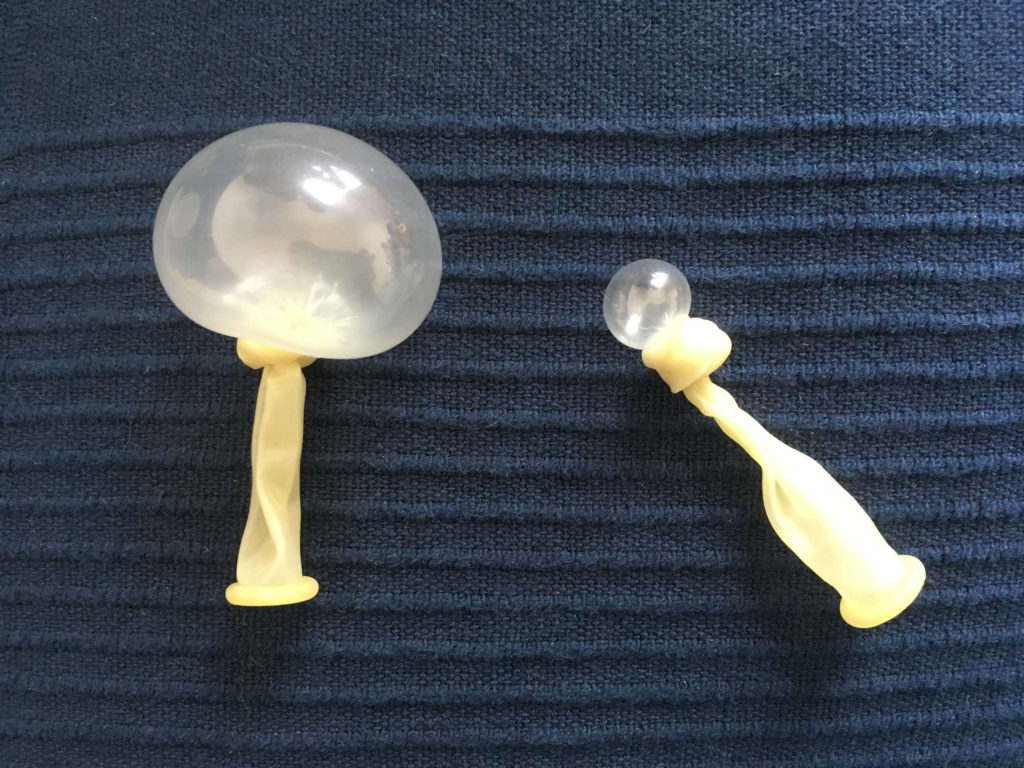
« On a aussi des choses comme ça… quand je vous disais que vous étiez un peu pauvres avec votre vessie de truite! »
« Là, tu vois le bilou! Il y a un, deux, trois muscles ; une vessie qui est tellement transformée qu’elle forme une structure squelettique! On a pu déterminer qu’il y avait un muscle qui se contractait après l’autre et à l’intérieur de la vessie; on a encore vu ça : trois membranes ! Et pour le moment on a pas encore compris à quoi servait cette structure. Ce poisson c’est la donzelle : l’ophidion rochei ».
« Mon idée, c’est qu’on a ici un bouchon, comme sur une bouteille de vin. Mais celui-ci ne va pas se détacher complètement, donc t’es capable de tirer dessus et il revient. A chaque retour, il y aurait le son. Le muscle qui est là, il va tirer vers l’arrière et là c’est directement relié au truc…. Comme un arc à flèches : au repos, tu as le bois et la corde et on tend le système. Ici, le bouchon est amené vers l’arrière et le muscle qui est là sert à tirer vers l’avant, mais comme il y a déjà une force vers l’arrière, quand le muscle se relâche, il revient beaucoup plus vite, évidemment ! Ce que je peux dire c’est que ce système change en fonction des individus, que les femelles n’en ont pas et qu’elles ne font pas le même son. »
Térence remarque que cela semble servir à accorder la résonance. Eric a essayé de comprendre ça avec le principe de résonateur de Helmholtz. Philippe explique que le résonateur de Helmholtz ouvert et non fermé comme ici. « oui, ça m’emmerde » dit Eric.
« Ma conclusion, bien que je ne comprenne pas, c’est que cette membrane doit aider à encaisser la variation de pression quand la boule revient. Comme si c’était une valve de pression. » Térence remarque qu’en fonction des fréquences, c’est comme si c’était ouvert le résonateur de Helmholtz : ça fait un filtre acoustique qui laisse passer les graves. Comme le déplacement peut être transmis c’est comme si c’était ouvert. Pour Philippe, la caractéristique le Helmholtz c’est de faire des résonances beaucoup plus graves que celles données par la taille d’un objet. On a pas besoin de caser une importante longueur d’onde dedans.
Celui-ci nous évoque un coquillage, il semble rigide. Cela expliquerait, selon Philippe, la présence de parois plus fines. Lia remarque qu’il y a quand même un lien avec notre modèle du « stick-slip » : c’est l’emmagasinement d’énergie. Les muscles soniques sont gros et occupe une place importante dans le corps. Le formol semble rendre les tissus tannés. Eric sort l’organe du formol. L’aspect est en effet plus celui du cartilage, comme le cuir, plutôt qu’une peau souple. C’est ce que nous avions observé chez le merlu provenant de notre poissonnier grenoblois.


Un troisième type de vessie présente un muscle à l’extérieur qui va tirer vers l’avant et à l’intérieur, un autre muscle replié. La vessie serait une extension du tube digestif et les muscles qu’on y retrouver sont donc un peu près les même (avec des contractions involontaires de 2,5 Hz seulement). Philippe remarque que le son produit par ces différents systèmes n’est pas fondamentalement différent, il s’agit toujours d’impulsions, bien que le timbre diffère.
Eric nous fait ensuite écouter deux poissons de taille différente qui produisent deux sons dissemblables. Nous regardons un autre exemple de deux poissons qui réalisent une danse pendant qu’ils produisent leurs sons (claquements de dents). Eric évoque aussi les plages horaires durant lesquelles seules certaines espèces de poissons chantent. Ainsi, différents chants peuvent être enregistrés à différents moment de la nuit.
A Tahiti, certains poissons émettent leurs chants dans une huitre. Celle-ci leur sert de haut-parleur pour émettre le son plus loin. On écoute ensuite le Térapon et le Pelates dont le son ressemble à un klaxon. Il est émis par des contractions très rapide de la vessie. Un autre poisson émet des son à 350Hz ce qui pour notre oreille donne l’impression d’un son continu. Les poissons n’entendent pas les bruits parasites (crevettes claquettes qui émettent à 700 Hz, par exemple) car son audition se limite à une certaine bande de fréquences.
A propos des fonctions de la vessie natatoire, certains poissons ont une vessie qui n’est pas collée mais placée à 30° de la colonne vertébrale. Elle se loge dans des os qui sont dans le prolongement de la nageoire. En plus, c’est comme si la poche se prolongeait en deux écouteurs qui viennent taper sur l’oreille. Ce poisson peut en outre aller se loger dans le sable et entendre le son des proies (les vers) qui se déplacent. La vessie transmet donc le son au système auditif. Térence remarque que c’est comme la réversibilité des systèmes micro/haut-parleur, un microphone dynamique peut-être employé comme haut-parleur, et réciproquement.
Certains poissons entendent aussi grâce au mouvement particulaire (pas seulement les ondes de pression). La vessie est capable de transformer l’onde de pression en mouvement particulaire. Autrement dit, un poisson qui possède une vessie natatoire entend mieux qu’un poisson qui n’en a pas. Et plus cette vessie est proche du crâne, meilleure est la capacité auditive du poisson. On en revient à la chaine de Weber évoquée précédemment : l’onde de pression impacte la vessie qui transmet cette vibration à une chaine d’osselets qui pousse le liquide auditif.
Les harengs sont capable d’entendre les dauphins grâce à une vessie qui se prolonge en tube, gros comme une aiguille d’épingle, et à l’autre extrémité on a une petite poche d’air à côté du cerveau. L’onde est donc transmise sans perte au cerveau. Ils ont aussi des « maculas » (cellules sensorielles) un peu plus développées que les autres poissons.
Dans le cerveau des poisson, on a de part et d’autre les oreilles qui sont composée de trois canaux semi-circulaires (pour l’équilibre) et en dessous les otolithes (cailloux d’oreille). Ces cailloux sont en relation avec une plaque recouverte de cellules ciliées qui n’ont pas toutes la même taille. En bougeant les cailloux, on fait bouger les cils qui sortent et un type d’influx nerveux est envoyé au cerveau. Si le son est plus fort, plus de cils bougent et un autre type d’influx nerveux est envoyé au cerveau. L’oreille et le crâne du poissons n’ont pas la même composition chimique et c’est ce décalage qui permet la transmission de l’influx. Si les « maculas » ne sont pas tout à fait pareil, le poisson est sensible à plusieurs types de variations dans l’oreille. La vessie est donc très importante pour l’audition : plus qu’une caisse de résonance, c’est une caisse d’amplification.
Philippe explique que l’impédance acoustique est un peu comme l’indice optique (réfléchie lorsqu’il y a un changement de milieu). Le passage du son de l’eau à l’air de la vessie va donc diminuer considérablement le signal : les ondes vont être réfléchies et ne vont pas passer dedans. Il y a un écho qui va être produit.
Philippe conclue qu’il a théorie : la production du son serait un petit choc élastique à un endroit de la vessie qui va se propager sur toute la surface et va être réémis. Tout cela dépend vraiment de l’élasticité de la membrane (Il ne faut pas que la vessie soit trop élastique.). Eric fait l’hypothèse qu’à partir d’un certain module de Young on serait obligé d’avoir une partie de la vessie plus mobile pour avoir de grandes amplitude. En dessous d’une certaine valeur, le muscle pourrait suffire, au dessus la création d’une vessie serait nécessaire.
Un poisson simple pour nos premiers tests serait le Tilapias (?) d’après Eric. Reste encore à résoudre la question de comment le muscle pourrait être simulé…