

L’après-midi débute avec une présentation des travaux de Thomas Duchenne, étudiant en master dans le laboratoire d’Eric Parmentier. Ses recherches consistent à stimuler une vessie natatoire avec un marteau piézo-électrique, qui mesure précisément la force avec laquelle il stimule la vessie. Il mesure ensuite le déplacement et la vitesse qui suit cette stimulation avec un vibromètre laser. Il réalise aussi des coupes histologiques pour analyser cela en microscopie optique pour voir si il y a différents paramètres qui permettent de caractériser la vessie.
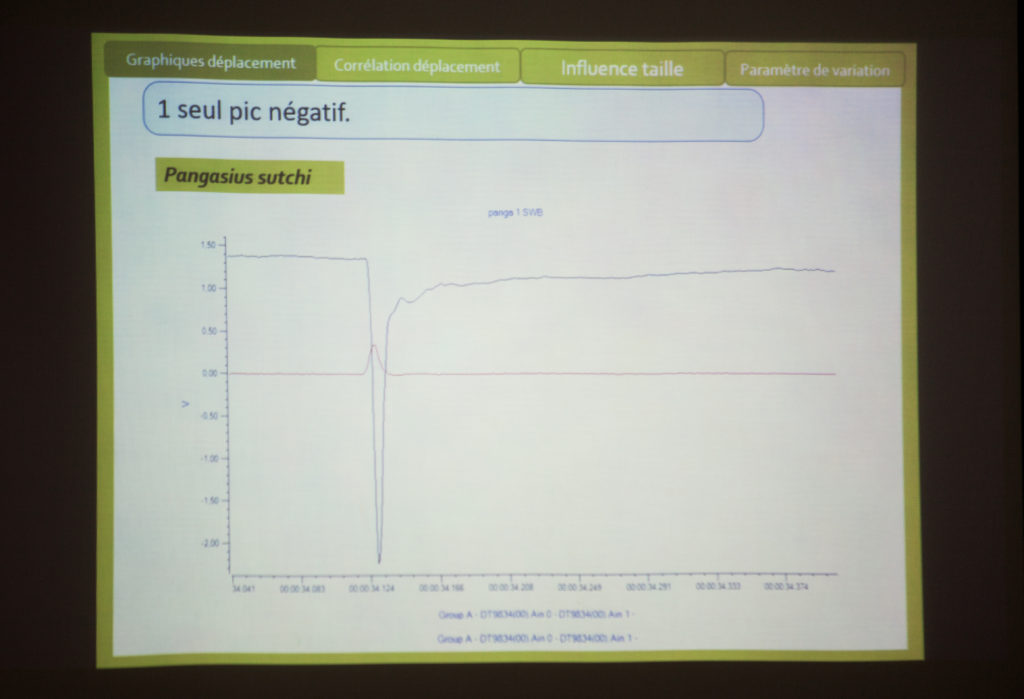
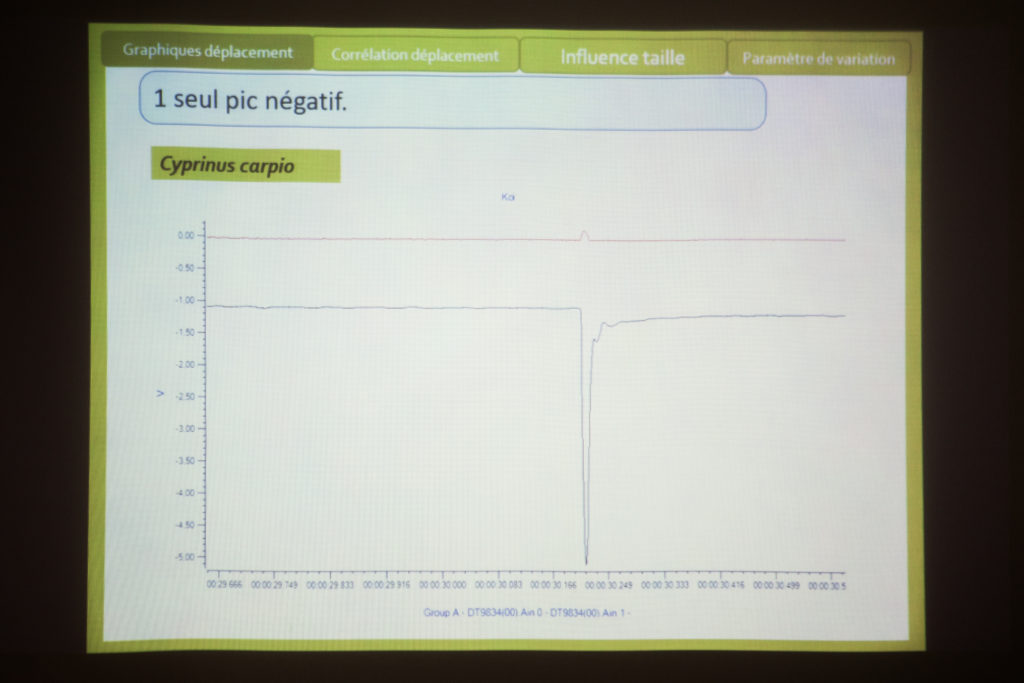
Pour le paramètre de déplacement : lorsqu’il stimule la vessie natatoire d’une Loche Brotia (type de poisson clown) avec le marteau, il obtient un pic puis la vessie descend et remonte. Elle ne résonne pas vraiment. Eric constate qu’il y a tout de même sur la courbe une petit oscillation et Philippe assure que cette courbe est caractéristique du comportement viscoélastique de la vessie. Le point où Thomas réalise sa mesure ne correspond pas exactement au point d’impact du marteau et cette réponse dure environ 50 ms. Chez le piranha à ventre rouge, Thomas observe plus ou moins la même chose, sans la petite oscillation. Chez le pangasus, il observe quelques petites oscillations qui reviennent lentement à un état d’immobilité. Ici encore, Philippe évoque les propriétés viscoélastiques du matériaux. L’éponge est par exemple un tel matériau : quand on la presse, elle revient lentement à son point d’immobilité. Pour Eric, cela serait donc lié à la composition de la vessie (collagène et viscosité du matériaux). Bien que petites, les oscillations correspondent selon Philippe à autre chose. Thomas trouve aussi ces oscillations chez la carpe koï.
L’appareil de Weber, qui existe notamment chez les poissons-chat, les piranha, les carpes, est une chaine d’osselets placée sur la colonne vertébrale qui relie la vessie natatoire au système auditif du poisson. Il intervient dans l’équilibre et les capacités auditives des poissons. Quand on appuie sur la vessie natatoire, les osselets bougent et augmentent les capacités auditives du poisson. Eric se demande si les petites oscillations pourraient être la réponse de ces petits osselets à l’impact du marteau, mais certains tests réalisés sur des poissons ayant un appareil de Weber ne présentent pas d’oscillations…
Philippe fait l’hypothèse que cette réponse est le résultat d’une onde circulant sur la paroi de la vessie à l’issue de l’impact avec le marteau. Il y aurait donc des vessies qui résonnent plus ou moins, mais Eric note que le signal obtenu ne repasse pas au dessus du point zéro. Thomas et Térence s’accordent pour dire que certains signaux sonores dont le point d’origine est déplacé peuvent rester en dessous du zéro.
Selon Philippe, la vessie n’est pas un oscillateur simple. Il serait intéressant pour lui de mesurer la réponse de la vessie à différents endroits pour savoir en quelle mesure la forme de la vessie intervient dans la propagation du signal. Philippe demande si les différences de signaux peuvent venir de la peau de la vessie, plus ou moins tendue. Pour Thomas, la forme n’est pas la même, mais la texture est cela assez équivalente, par exemple entre la carpe et le poisson-chat.
Lia s’interroge sur les différences de formes entre les deux espèces. Chez la carpe, la chambre antérieure est plus grande que la chambre postérieure. Dans ses expériences, Thomas a tapé sur la chambre antérieure car la postérieure était trop petite. Chez le poisson-chat, les deux chambres sont séparées par une sorte de « pilier » et là encore, Thomas a du se placer sur la chambre antérieure pour ne pas toucher le pilier et parce que la chambre postérieure était trop petite. Les formes varient finalement beaucoup et Thomas doit donc parfois déplacer sa mesure d’un centimètre.
Philippe demande si la vessie se dégonfle durant les expériences. Thomas ferme le canal pour éviter que l’air ne s’échappe, mais il est vrai que l’aspect change un peu, notamment car le corps du poisson sèche à l’air libre et la vessie devient légèrement transparente… Thomas observe encore ces oscillations chez les cichlidés mais la cage thoraciques est ici accrochée au corps de l’animal. Eric en a discuté avec M. Fine : dans ce cas précis, la résonnance ne serait pas celle de la vessie natatoire, mais de l’ensemble de la cage thoracique. D’après Michael Fine, ouvrir le poisson empêcherait un amortissement de la vessie dû aux tissus environnants. S’il y avait vraiment une résonance, on pourrait l’observer en tapant dessus. Thomas a fait le test avec une membrane de ballons et a observé que les plages d’oscillations n’étaient pas vraiment comparables à ce qu’il observe sur les vessies. Cependant il n’a pas essayé sur un ballon gonflé.
Thomas observe aussi que le déplacement dépend de la force d’impact. Grâce à ce rapport force/déplacement, Thomas à pu comparer différentes espèces et montrer que ces réponses ne sont pas les même selon les espèces : par exemple la réponse est nettement plus forte chez le piranha. Une autre façon de faire est d’observer la pente de la régression linéaire chez les différents individus. Après analyse statistique, Thomas n’a pas pu obtenir de courbes significatives car les population sont trop variables (parfois un individu, parfois cinq). Dans une publication de M. Fine, la courbe de réaction (déplacement) après stimulation du muscle sonique, varie selon la taille de l’animal. C’est ce que Thomas souhaiterait montrer par son approche statistique.
Il a réalisé le même type d’expérience pour observer la vitesse. Il observe par ce moyen des réaction assez similaires que dans son étude du déplacement. Chez certains poissons il observe plusieurs cycles de réaction. Ce qui étonne Thomas c’est que pour plusieurs types de ciconidés (dont la vessie est assez similaire), c’est juste la taille qui semblerait justifier une pente différente.
Plusieurs facteurs semblent influer sur les propriétés vibratoires de la vessie : la taille de la vessie, la pression en gaz à l’intérieur de la vessie (lorsque les poissons sont euthanasiés il est possible qu’ils relâchent une partie de leur air), la zone impactée par le marteau (il est difficile pour Thomas d’être précis avec des structures de vessies aussi variables).

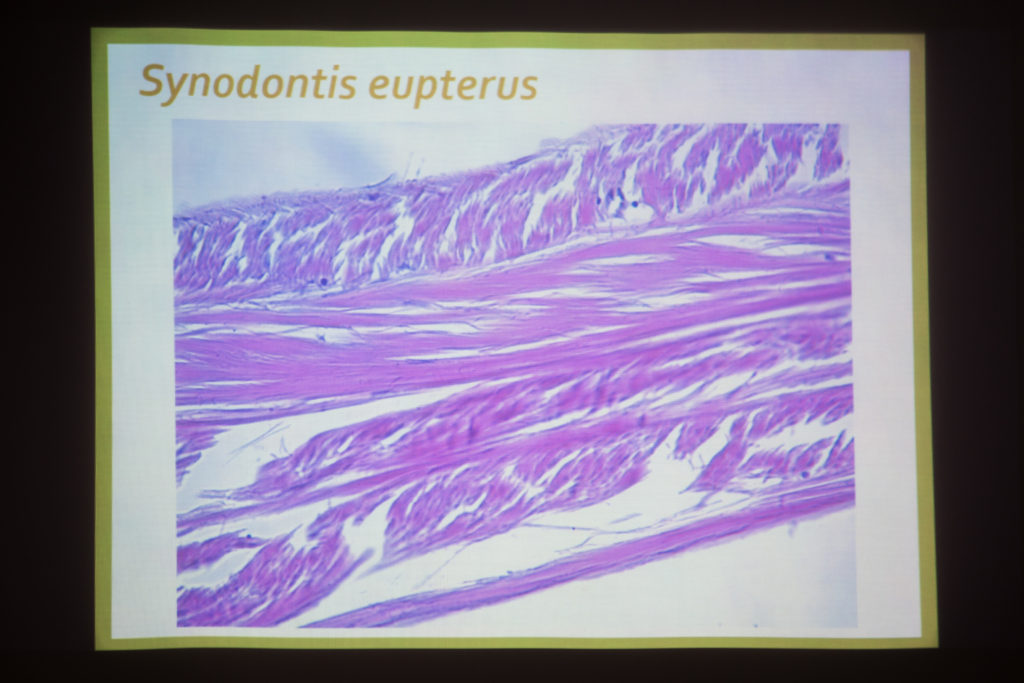
Thomas nous montre ensuite les coupes histologiques. L’orientation des fibres varie si la coupe est réalisée sur la partie dorsale ou latérale. Thomas n’a pas mesuré l’épaisseur, notamment car le formol modifie les tissus. Il note aussi qu’entre les différents modes de fixation utilisés, les tissus sont plus ou moins altérés (le formol altère plus que le Boia (?)). Sur les images, on observe bien les différentes couches de fibres.

Comme Philippe le remarque, Thomas n’a pas mesuré et comparé la durée de l’oscillation au début de ses tests. Il a pu néanmoins observer plus tard que cette durée varie selon les espèces et en fonction de la force de frappe. Pour le moment il n’a pas encore assez de données sur ce point. Pour Philippe, la fréquence d’oscillation et le temps de retour à la normale sont deux mesures intéressantes. Chez la carpe koï, la force de frappe ne semble pas avoir d’influence sur la période de réaction de la vessie tandis que chez d’autres espèces c’est le cas. D’après Eric, la remarque de Philippe sur une possible répercussion de l’impact entre les deux chambres de la vessie est intéressante.
Philippe demande ce qui détermine la pression de gaz dans la vessie. Pour Thomas, il y a deux types de vessie : la première possède un canal pneumatique qui fourni du gaz; les espèces qui n’ont pas de canal ont une glande à gaz. Il y a toujours du gaz et une mise en tension dans la vessie. Le poisson module ce volume de gaz. Le poisson a flottabilité neutre, dépendante d’un certain volume de gaz, uniquement à une profondeur donnée. Quand il remonte ou descend, il change cette flottabilité (comme le plongeur qui expulse l’air en remontant). D’après Eric, lorsque le poisson remonte, le gaz et réabsorbé et passe dans le système sanguin. Si cette air n’est pas expulsé lors de la remontée, le poisson explose. La vessie est tendue mais ne change pas énormément de volume. Dans le poisson, cette poche est tout de même contrainte par ce qui l’entoure. Comme pour nous les côtes sont libres mais limitent tout de même son expansion.
Pour Philippe, le module de Young pourrait montrer la résonance de la poche de gaz dans la mesure où il prend en compte la nature de l’enveloppe (plus l’enveloppe est rigide, plus elle résonnerait à haute fréquence). Pour Éric il pourrait être intéressant d’avoir cette donnée. Avec le module de Young on mesure le point de saturation d’une élasticité lorsqu’on étiré un matériau dans un certain sens. D’après Eric, le travail de Thomas est intéressant car il s’y distingue deux types d’élasticité des vessies. Lia note que l’élasticité dans le sens latéral et longitudinal n’est pas le même. C’est ce que présument aussi les images en microscopie optique où ces deux structures des tissus diffèrent.